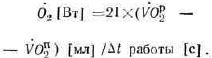При любом использовании данного материала ссылка на первоисточник обязательна!
педа от скорости вращения колес. При увеличении вращения колес увеличивается и нагрузка на педали, которая возрастает от 28,5 до 54 кгс при скорости от 28,5 до 73,1 км/ч.
Для создания «наката» велосипеда на одном из роликов велостанка, лучше на 2-м или 3-м, устанавливается диск с массой 5 кг. Чтобы не было биения, диск с массой необходимо сцентровать с роликом.
Настоящее тормозное устройство просто по конструкции, портативно, удобно в эксплуатации. Оно может быть установлено на велостанках различных конструкций. Применение его обеспечивает возможность регулировать нагрузку на велостанке в широком диапазоне, что способствует более эффективной подготовке спортсменов в осенне-зимнем периоде тренировки.
НАУКА — СПОРТУ
О. И. Федоткина, А. И. Головачев,
А. Ю. Колесников, Ю. Г. Крылатых, ГЦОЛИФК, ВНИИФК
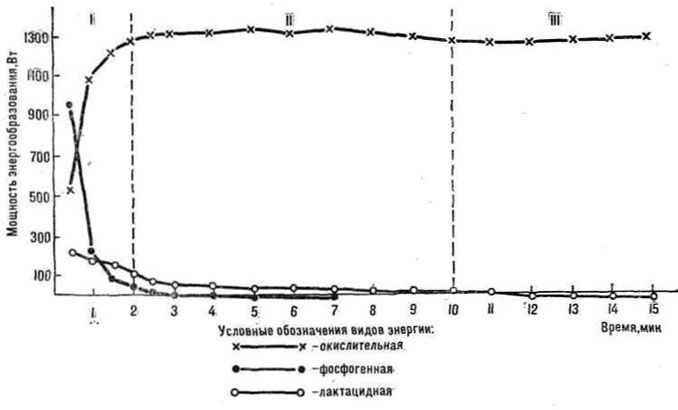
Известно, что интенсивная мышечная работа обеспечивается суммарным вкладом трех основных энергетических систем: окислительной, фосфогенной и лактацидиой (Я. М. Коц, 1982). Каждая из них характеризуется мощностью, емкостью и эффективностью (Н. И. Волков, 1974). Лимитирующим фактором образования энергии окислительным путем а начале выполнения упражнения является его инерционность. Дефицит кислорода в начале работы покрывается за счет образования энергии в анаэробных процессах (фосфогенного и лактацидного). Однако при определении динамики энергетических затрат исследователи в основном использовали измерение скорости потребления кислорода во время работы в лишь в отдельных случаях учитывали скорость образования энергии гликолитическим путем (Wasserman К., van Kessel A. Z., Burtong, 1967). Мощность фосфогенной энергетической системы, как правило, измеряли по специальному тесту Margaria R. (1968).
В последние годы на основе экспериментальных работ Fox (1974) стало возможным рассчитать мощность фосфогенного энергообразования при кратковременной предельной работе по алактатной фракции кислородного долга. Такой подход проведения расчета в предельных упражнениях большой относительной мощности не применялся, и суммарные энергетические затраты рассчитывали без вклада фосфогенной системы.
В связи с этим целью нашей работы
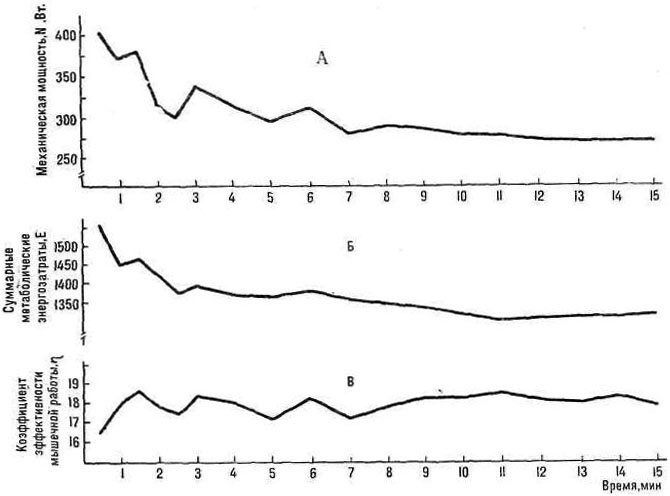
явилось изучение динамики суммарных энергетических затрат во время выполнения предельной мышечной работы относительной мощности с учетом трех основных энергетических систем.
Задачи исследования:
1. Определить изменения скорости образования энергии окислительным, лактацидным и фосфогенным путем.
2. Определить динамику коэффициента эффективности мышечной работы во время выполнения упражнения.
Для решения поставленных задач были проведены экспериментальные исследования, в которых участвовали 4 спортсмена I разряда в возрасте 16 лет. Перед началом исследований все спортсмены проходили тестирование функциональной подготовленности на основе выполнения нагрузки ступенчато-нарастающей мощности до отказа. Характеристика испытуемых и результаты предварительного исследования представлены в табл. 1.
При проведении основного исследования каждому спортсмену предлагалось выполнить предельную нагрузку при произвольном распределении усилий в течение 15 мин с целью достижения максимальной производительности. Спортсмены выполняли это упражнение 13 раз, однако до конца использовалась только одна нагрузка. Во всех остальных случаях работа прерывалась через неизвестное для испытуемого время от начала упражнения. Остановки проводились через 30 с, 1, 1,5, 2, 2,5, 3, 4, 5, 7, 8, 10,